
Antes de producirse la fecundación, el espermatozoide, debe atravesar la corona radiada y la zona pelúcida que recubren y protegen al oocito, para poder fusionarse con su membrana.
1-Penetración de la corona radiada:
De los 200-300 millones de espermatozoides de media que contiene un eyaculado, solo unos 300-500 llegan al lugar donde se produce la fecundación, una de las teorías posibles es que el resto de espermatozoides que quedan en el camino sirvan para apoyar al resto.
La corona radiada puede tener varias funciones en la fecundación:
-Al hacer más voluminoso al oocito, facilita el encuentro con los espermatozoides.
-Atraer a los espermatozoides que ya realizaron la reacción acrosómica.
-Aumenta la vida fértil del oocito evitando que se endurezca la zona pelúcida (ZP).
-Facilitar la llegada del espermatozoide a la ZP.
-Filtrar la llegada de espermatozoides no aptos.
Esta corona radiada está formada por células de la granulosa unidas por una matriz de ácido hialurónico que se va expandiendo tras la ovulación.
Al llegar a la corona radiada, solo aquellos espermatozoides capacitados y con el acrosoma intacto pueden penetrarla, viendo modificado su movimiento debido a la viscosidad que presenta la estructura, que ofrece resistencia a la penetración espermática. La interacción con el ácido hialurónico, propicia un movimiento más lineal y progresivo, por lo que se distinguen un componente mecánico y otro enzimático que facilitan la penetración de la corona radiada.
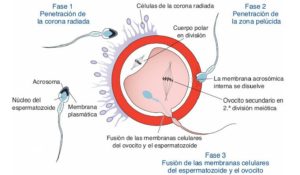
2-Penetración de la Zona Pelúcida:
La ZP es una cobertura glicoproteica que rodea al oocito y presenta varias funciones:
-Es especie-específica, es decir, facilita que espermatozoides de la misma especie la atraviesen, dificultándolo para los de otras especies.
-Induce la reacción acrosómica en el espermatozoide.
-Evita que el oocito sea fecundado por más de un espermatozoide.
-Evita que las blastómeras que forman el embrión antes de implantar se disgregen o implanten en lugares ectópicos.
La estructura de la ZP es diferente en cada especie, aunque en la mayoría se pueden diferenciar dos capas. Una más interna y densa, la otra, externa y más laxa.
Se pueden diferenciar 2 etapas diferentes para describir cómo penetra el espermatozoide la ZP:
1-Fijación primaria: el espermatozoide se une a la ZP con el acrosoma intacto.
2-Fijación secundaria: la unión se da tras completar la reacción acrosómica.
La ZP de la especie humana al igual que la de ratón y hámster, esta formada por 3 familias de glicoproteínas llamadas ZP1, ZP2 y ZP3. Cada una de ellas tiene diferentes funciones que explicamos a continuación:
-ZP1: su función parece ser meramente estructural, se ha observado en ZP sin ZP1 que se produce la fertilización tanto in vivo como in vitro.
-ZP2: se unen a la membrana acrosomal interna, es decir, la que queda expuesta cuando se ha completado la reacción acrosómica.
-ZP3: se une a la membrana externa del espermatozoide, creando la primera unión. Además, se diferencian sitios con diferente afinidad de unión, y aquellos con menor afinidad, parecen ser los encargados de desencadenar la reacción acrosómica.
Una vez conocemos todos los actores que participan en el mecanismo de penetración de la ZP, vamos a explicar el proceso de forma más detallada.
La membrana plasmática del espermatozoide debe contener algún ligando que se une en la denominada fijación primaria a la ZP, interactuando con ZP3. Este ZP3, propicia además la reacción acrosómica, favoreciendo dos procesos, la unión entre ZP2 y membrana acrosomal interna, fijación secundaria, y la liberación de la acrosina y proacrosina, que actuarán permitiendo el avance del espermatozoide por la ZP.
De esta forma, la proacrosina liberada se activa y transforma en alfa-acrosina, capaz de hidrolizar la ZP, así el espermatozoide puede avanzar. Esta alfa-acrosina, se autocataliza en beta-acrosina, volviendo a hidrolizar la zona y degradándose posteriormente. De esta forma cíclica, el espermatozoide consigue avanzar por la ZP hasta llegar a la membrana del oocito.
El espermatozoide tarda pocos minutos en atravesar la ZP, por lo que es indispensable que existan mecanismos de control para evitar una polispermia, la fecundación del oocito por más de un espermatozoide. Se diferencian dos tipos de bloqueos:
-Bloqueo primario: solo se da en conejos. Se produce una rápida despolarización de la membrana del oocito, que impide la fusión de otro espermatozoide.
-Bloqueo secundario: se produce una modificación estructural de la ZP. Se origina por la reacción cortical producida tras la fusión de un espermatozoide. En ella, se liberan enzimas presentes en los gránulos del citoplasma al espacio perivitelino, propiciando el endurecimiento de la ZP. Este proceso, está regido por el flujo de calcio al interior del oocito.
3-Fusión de membranas espermatozoide-oocito:
Tras atravesar la corona radiada y el ZP, el espermatozoide llega al espacio perivitelino y al contactar con la membrana del oocito, este se recuesta sobre ella.
El primer punto de unión se origina en la zona ecuatorial de la cabeza del espermatozoide. La unión se produce gracias a la interacción de integrinas presentes en la membrana del oocito y sus ligandos, desintegrinas, distribuidas por la membrana del espermatozoide. Tras esta adhesión, se produce la fusión, que continúa hacia la zona posterior de la cabeza y zona intermedia del espermatozoide, no fusionándose nunca la membrana acrosómica interna.
Se han descrito otras proteínas que actúan en la fusión de las membranas, como por ejemplo la epididimaria DE, proteínas de la familia ADAM y proteína transmembrana CD9.
Esta fusión entre gametos, se puede originar en cualquier parte de la membrana del oocito, a excepción de una zona que no tiene microvellosidades, que parecen contener los receptores para los espermatozoides. Esta zona se encuentra sobre el huso meiótico y es por donde se produce la extrusión del primer corpúsculo polar.
El contacto entre las membranas de los gametos, origina la liberación de gránulos corticales del oocito, que por la acción de enzimas lisosómicas, evita que otros espermatozoides entren al interior del oocito, originando una poliespermia.
A diferencia de lo que se creía, el espermatozoide entra completamente al interior del oocito, eso sí, dejando su membrana plasmática en el exterior.

Una vez dentro, se produce la división entre la cabeza y la cola del espermatozoide. Además, la mayoría de organelas del espermatozoide degeneran, como por ejemplo sucede con las mitocondrias, que siempre son de herencia materna. Si se conserva el centriolo espermático, que originará el áster, esencial para el movimiento de los pronúcleos. Por último, se rompe la envoltura nuclear y se descondensan los cromosomas espermáticos.
En próximas entregas, continuaremos explicando el proceso de la fecundación, que aún no ha acabado aquí, mientras tanto, puedes leer entregas anteriores en nuestro blog:
Bibliografía:
- Langman. Embriología Médica, 13ª ed. Barcelona, España: Ed.Lippincott/Williams & Wilkins, 2016.
- Infertilidad «Fisiología, diagnóstico y tratamiento». Gustavo Pagés ; Juan Aller. 2006.
- The miracle of life. Lennart Nilsson, 1980.